














O šíření signálů v živých organizmech
Kdo měl tolik trpělivosti, že se prokousal všemi předchozími částmi, ten snad už získal jakousi představu o tom, jak složité a různorodé jsou signály, které bez ustání ve dne v noci putují všemi směry naším složitým a rozvětveným nervovým systémem. Přenos a zpracování signálů nervovým systémem byl dlouho považován za základní a nejdůležitější mechanizmus, kterým se mezi sebou dorovzumívají jak jednotlivé buňky, tak různé orgány. Víme, že řídí, reguluje a kovntroluje nejen naše pohyby, reakce a stavy mysli, tedy veličiny, které si uvědomujeme, ale i ty, které se dějí bez našeho vědomého přispění, jako např. srdeční činnost nebo trávení. Víme též kam vede, jak se jmenuje a k čemu slouží každý nepatrný kousíček nervu. Dlouho jsme si mysleli, že porozumět činnosti centrálního a periferního nervového systému stačí k vysvětlení většiny toho, co se v nás děje. V posledních letech byl objeven a popsán jiný způsob přenosu signálů, o kterém však zatím dobře nevíme, k čemu všemu by mohl sloužit. Tento způsob signalizace není založen na změnách elektrického potenciálu, jako je tomu v nervovém systému, ale na změnách vnitrobuněčné koncentrace vápníku.
Regulace intracelulárního vápníku
Biologové zjistili již dávno, že si buňky udržují ve svém vnitřním prostředí nesmírně nízkou koncentraci volného vápníku. V normálních podmínkách je to asi kolem 50 nM (připomeňme, že n je zkratka pro nano, značící tisícinu miliontiny). Bylo to překvapující, protože jeho extracelulární koncentrace je řádově 2 – 5 mM, což je téměř stotisíckrát více. Tento chemický gradient reprezentuje ohromnou sílu, která nutí vápník vrátit se do buňky. Navíc, jak si možná pamatujete z první části, je vnitřní prostředí elektricky negativní vzhledem k prostředí vnějšímu. A protože vápník je kationt, a k tomu dokonce dvojmocný (Ca2+), snaží se také elektrické síly vtáhnout ho dovnitř. Naštěstí vápníkové kanály, kterých je v buněčné membráně několik druhů, jsou při normálních membránových potenciálech velice málo propustné. Ale pozor, většina z nich jsou napěťově závislé a při snížení hodnoty membránového potenciálu se otevírají, obdobně jako draselné a sodíkové kanály, které jsou odpovědné za vznik akčních potenciálů (viz Vesmír 75, 68, 1996/2).Vápníku je v buňkách celkově daleko víc, jenže je tam různě poschováván. Částečně je navázán na některé intracelulární molekuly, ale hlavně, a to je pro další výklad důležité, je uskladněn ve dvou tajuplných skrýších, do kterých je neustále přepumpováván. Z intracelulárního prostoru je tedy vápník odčerpáván třemi směry. Zaprvé přes plazmatickou membránu z buňky ven. Zčásti se o to stará vápníková pumpa, která k tomu získává energii spalováním kyseliny adenozintrifosforečné (ATP). Jenže sama na to nestačí. A tak jí nepřímo pomáhá sodíko-draslíková pumpa. Jak to, vždyť ta se má starat o udržování gradientu draslíkového a sodíkového! Jenže v buňečné membráně existuje řada mechanizmů, které na sodíko-draslíkové pumpě parazitují. A to tak, že využívají energie uskladněné v sodíkovém gradientu, aby pomohly jiným iontům nebo látkám k opuštění buňky. Jedním z nich je právě vápník. Sodík díky svému gradientu má tendenci do buňky vstupovat. Toho vápník využívá tak, že ho tam pustí jen pod podmínkou, že si s ním vymění místo. Tento druh transportního mechanizmu, založeného na výměně jedné molekuly za jinou, je v přírodě poměrně častý a připomíná trochu hru škatule, škatule, hejbejte se. V případě sodíko-vápníkového výměníku vstupují do buňky tři sodíky, a zároveň umožní jednomu vápníku buňku opustit. (Ve skutečnosti je to o něco složitější, protože té výměny se zúčastňuje též draslík.)
K udržení a regulaci tak nízké intracelulární koncentrace si buňka pomáhá také tím, že si velkou část vápníku převádí vápníkovou pumpou, podobnou té, která je v buněčné membráně, do jakýchsi zvláštních skladů. Z obou může být vápník v případě potřeby vypuštěn do vnitrobuněčného prostředí. Rozdíl mezi nimi spočívá v mechanizmech, kterými se vyprazdňují. Vrátka jednoho z nich se otevírají pod vlivem zvýšené hladiny 1,4,5-trifosfátu (IP3). Ten vzniká z fosfatidylinositol l-4,5-bifosfátu (PIP2) působením fosfolipázy C. Vrátka druhého skladu mají zámek založený na jiném principu. Jedním z klíčů, které zámek otevírají, je kofein. Často se používá při různých experimentálních strategiích, chceme-li nějak tento sklad násilně vyprázdnit. Fyziologickým stimulem však je, a to celkem překvapivě, právě samotný vápník. Jeho nízká koncentrace udržuje vápník uzavřený ve skladu, překročíme-li však určitou prahovou koncentraci, otevřou se vrátka dokořán a vápník se vyhrne ven. Delší působení vápníku nebo jeho příliš vysoká koncentrace však dvířka opět přibouchne. Mechanizmů, které se na regulaci hladiny intracelulárního vápníku podílejí, je mnoho (viz obrázek).
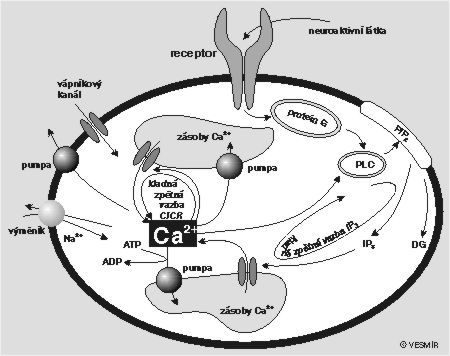{kind=link}
Jak měřit změny intracelulárního vápníku
Intracelulární vápník je jakousi centrální veličinou, na které závisí téměř všechny buněčné regulační mechanizmy. Volného intracelulárního vápníku je v buňkách nepatrně, a proto stačí přidat (nebo ubrat) jen mizivé množství, aby to vyvolalo okamžité a hluboké změny v jeho koncentraci. O tom se vědělo již dávno. Problém byl však v tom, že experimentální sledování změn tak nízkých vápníkových koncentrací není zrovna jednoduché. V 80. letech vyvinul Robert Tsien fluorescentní látky, zejména fura-2 a fluo-3, které mění své optické vlastnosti v závislosti na přítomnosti i nepatrných množství vápníku. Již dlouho předtím se k témuž účelu používalo jiné látky, ekvorinu. Nové látky přinesly pokrok ve dvou směrech. Zaprvé je není třeba do buňek složitě vpravovat. Před použitím se esterifikují, což jim dodá schopnost snadno prostupovat lipoidními membránami. Jakmile se takto dostanou do buňky, vrhnou se na ně intracelulární esterázy, látka je hydrolyzována a tím ztratí svoji lipofilní vlastnost. Zůstane tedy uvězněna v buňce jako v kleci. Ta druhá výhoda spočívá v tom, že jejich fluorescence je mnohonásobně vyšší, než byla fluorescence ekvorinu, takže pro úspěšný pokus stačí, aby byly v buňkách přítomny v celkem rozumných koncentracích. Prakticky probíhá sledování hladiny intracelulárního vápníku tak, že po inkubaci s některým z výše uvedených optických indikátorů jsou buňky umístěny pod speciálně upravený mikroskop, kde jsou střídavě osvětlovány dvěma vlnovými délkami. Na obě vlnové délky odpovídá indikátor fluorescencí (s maximem obvykle kolem 540 nm). Vtip je v tom, že na jednu vlnovou délku (většinou kolem 340 nm, záleží na typu fluorescenční látky) odpovídá hlavně ta část látky, která vytvořila s intracelulárním vápníkem komplex, při druhé (obvykle 380 nm) odpoví zbytek, tedy ta část, která zůstala volná. Změny hladiny vápníku poruší rovnovážný stav mezi látkou, která vytvořila s vápníkem komplex, a tou, která zůstala volná, a tím se také mění měřená fluorescence. Tato metoda dnes umožňuje sledování změn vápníku během různých experimentálních postupů.Vápníkové oscilace
O vápníkových oscilacích v astrocytech jsme se již zmínili v předchozí části. Za poslední léta však byly popsány i v několika jiných typech buňek a je pravděpodobné, že budou objeveny v mnoha dalších.Vápníkové oscilace jsou vlastně sledem prudkých změn, jakýchsi špiček v koncentraci intracelulárního vápníku. Jak taková koncentrační špička vlastně může vzniknout? Porovnáme-li její tvar s tvarem akčního potenciálu, zjistíme, že jsou si oba průběhy podobné a liší se pouze v časové základně. Akční potenciál trvá něco kolem milisekundy, špička vápníkové koncentrace asi tisíckrát déle. Porovnejme tedy mechanizmy, které vedou ke vzniku těchto jevů. Akční potenciál je generován tím, že nějaká depolarizace nervové buňky otevře napěťově závislé sodíkové kanály, sodík začne proudit do buňky a tento proud kladných elektrických nábojů způsobí další snížení membránového potenciálu a tím ještě větší otevření sodíkových kanálů, a tudíž ještě větší proud. To je příklad klasické kladné zpětné vazby, která, není-li kompenzována jinou, silnější, a hlavně zápornou zpětnou vazbou, působí katastrofy různého druhu. Jako příklad mohou sloužit všemožné konflikty na poli rodinném (manžel dostane vynadáno, že přišel pozdě domů, příště tedy přijde ještě později, protože co by doma dělal, je-li mu tam jen spíláno) i mezinárodním (třeba nedávné závody ve zbrojení). S tím sodíkem to však tak daleko nedojde, sodíkové kanály se samovolně uzavřou, místo nich se otevřou kanály draslíkové a to vše způsobí, že se potenciál membrány navrátí ke své klidové hodnotě. S vápníkovými oscilacemi je to podobné.
Pokusme se nyní pomocí obr. na s. 336 sami nalézt nějakou kladnou zpětnou vazbu, která by mohla být příčinou prudkého nárůstu vápníkové koncentrace. Výchozím bodem pro nás musí být receptor, nacházející se v buněčné membráně a přístupný určité aktivní látce (neurotransmiter, hormon aj.) přicházející z vnějšku. Vnější podnět způsobí jeho aktivaci, a ta způsobí, že se přes G-protein a fosfolipázu C (PLC) začne „vyrábět“ IP3. Tady už tu kladnou zpětnou vazbu začínáme tušit. Víme totiž, že zvýšená hladina IP3 otevírá vrátka jedné z vápníkových rezerv. To způsobí, že se vápník vyhrne ven a jeho koncentrace v buňce začne stoupat. Jak známo, fosfolipáza C je aktivována vápníkem. Čím víc vápníku, tím víc IP3 a čím víc IP3, tím rychleji se vyplavuje vápník. Na této kladné zpětné vazbě je založena jedna z teorií vysvětlujících vznik vápníkových koncentračních špiček. Jenže tato zpětná vazba není jediná a čtenář jistě vysleduje z obrázku i jinou, založenou na tom, že dvířka druhého rezervoáru se otevírají při zvýšené hladině intracelulárního vápníku (čím víc vápníku v buňce, tím víc se ho uvolňuje ze skladu). Toto vyplavování se pravděpodobně děje pod vlivem cyklické adenozindifosfátribózy (viz též Vesmír 74, 689, 1995/12). Tato druhá kladná zpětná vazba celou problematiku komplikuje a dnes nikdo přesně neví, která z těch dvou je důležitější. Teorií je hodně, průkazných experimentálních ověření méně.
Vápníkové signály přenášejí frekvenčně modulované informace
Změny ve vápníkové koncentraci slouží k přenosu informace. Z obecného hlediska je při každém informačním přenosu důležité, aby byl ze zpracovávaného signálu vyloučen šum. Podíváme-li se blíže na vápníkové oscilace, zjistíme, že amplitudy jednotlivých špiček dosahují hodnot několikanásobně vyšších, než je základní hodnota „klidové“ vápníkové koncentrace. Tím je v podstatě podmínka vyloučení šumu splněna.Druhým důležitým faktorem je detekce a dešifrování přeneseného signálu. Radioinženýři dobře vědí, že zvukové (a samozřejmě i všechny ostatní) signály se přenášejí daleko věrněji, jsou-li frekvenčně modulovány, než když jsou modulovány amplitudově. Detektor (nebo demodulátor), který umí spočítat, kolik došlo signálů určitého typu (např. vápníkových špiček), pracuje obvykle se stoprocentní přesností. Naproti tomu detektor, který má změřit amplitudu přenášeného signálu, nám dodá výsledek ve formě čísla s konečným počtem desetinných míst. Digitální způsob záznamu a zpracování informace je daleko přesnější, a proto se ho dnes používá již běžně. Setkáváme se s ním každodenně ve styku s počítači nebo při přehrávání kompaktních desek. Je zajímavé, že na co přišli lidé po desítkách let bádání, tím se příroda řídí od nepaměti. Zdá se nám to překvapivé, protože se odjakživa většinou dorozumíváme sděleními modulovanými amplitudově. Máme-li bolesti v noze, říkáme, že nás tady někde něco bolí. Nenapadlo by nás podat tuto informaci digitálně – říci třeba, že z místa o souřadnicích 369 na 636 přichází signál typu bolest o intenzitě 963. A přitom do našeho mozku přichází něco, co se právě tomuto typu informace podobá. Žádné „tam někde uprostřed lýtka“! 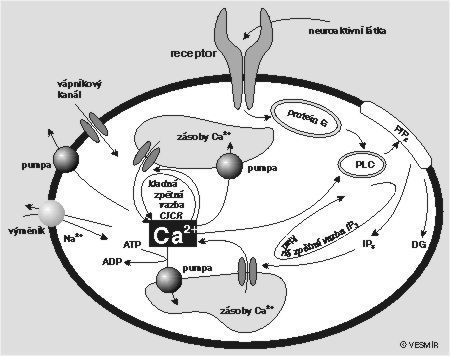 Podle nervových vláken, která informaci přinesla, identifikuje náš mozek přesně bod, kde je umístěno čidlo pro bolest, na které je napojeno příslušné nervové vlákno. Podle počtu vzruchů (akčních potenciálů) za jednotku času, tedy podle jejich frekvence, pozná, jak silná tato bolest je (opět zjednodušujeme, ale ne moc). Vápníkové oscilace jsou též jedním z příkladů frekvenčně modulovaného biologického signálu. Mohou být vyvolány řadou podnětů, ale většinou to bývá aktivace receptoru příslušnou látkou. Zvyšováním koncentrace této látky (na př. vasopressinu, adrenalinu, acetylcholinu, angiotenzinu aj.) se tvar vápníkových špiček téměř nemění, avšak zvýší se jejich frekvence.
Podle nervových vláken, která informaci přinesla, identifikuje náš mozek přesně bod, kde je umístěno čidlo pro bolest, na které je napojeno příslušné nervové vlákno. Podle počtu vzruchů (akčních potenciálů) za jednotku času, tedy podle jejich frekvence, pozná, jak silná tato bolest je (opět zjednodušujeme, ale ne moc). Vápníkové oscilace jsou též jedním z příkladů frekvenčně modulovaného biologického signálu. Mohou být vyvolány řadou podnětů, ale většinou to bývá aktivace receptoru příslušnou látkou. Zvyšováním koncentrace této látky (na př. vasopressinu, adrenalinu, acetylcholinu, angiotenzinu aj.) se tvar vápníkových špiček téměř nemění, avšak zvýší se jejich frekvence.
Přijměme tedy fakt, že buňky jsou schopny frekvenčně modulovat důležité informace. Jenže umějí je také demodulovat? Existuje nějaký biologický dekodér těchto informací, předávaných digitální formou? Odpověď zní: samozřejmě ano. Náš centrální nervový systém je právě takovým dokonalým dekódovacím zařízením, které je schopno informace rozluštit, zpracovat a předat na místo určení. Je jisté, že i dekodér vápníkových špiček existuje, jenže toho o něm víme málo. Zatím byly formulovány jen hypotézy založené na principu vápníkem stimulované fosforylace a defosforylace proteinů.
Vápníkové vlny
Vápníkové oscilace se šíří uvnitř jediné buňky a leckdo by mohl nabýt dojmu, že to vlastně žádný přenos informace není. Avšak nezapomínejme, že i na úrovni jediné buňky je třeba posílat informace z jednoho jejího místa na jiné. A hlavně, ve skutečnosti vůbec nejde jen o tu jednu buňku. První buňka slouží jen k jakémusi „nastartování“ procesu. Rozběhne-li se tento mechanizmus v jedné buňce, je velká pravděpodobnost, že se „rozezvučí“ jedna po druhé i buňky okolní. Příkladů je bezpočet. Omezíme se na buňky gliové. Jednak proto, že jsou svým umístěním nejblíže buňkám nervovým a tak mohou odposlouchávat a přebírat signály procházející nervovým systémem, že je mohou pozměňovat a modulovat, ale také na ně reagovat a odpovídat 1) , 2) , 3) . A pak také proto, že jsme si o nich v minulých částech už leccos řekli. Mluvili jsme zde o vápníkových oscilacích, které v astrocytech viděla a popsala jako první Ann Cornellová-Bellová. Neřekli jsme ale, že vzápětí po buňce, ve které oscilace vznikly, se začne měnit vápníková koncentrace i v buňkách sousedních 4) . V těch však nedochází přímo k vápníkovým oscilacím. Jde spíš o zvýšení hladiny vápníku, které trvá několik vteřin a pomalu se přenáší do sousedních buněk. Tím vzniká jakási vápníková vlna, která se šíří rychlostí 10 – 50 mikrometrů za sekundu od buňky k buňce. Její amplituda, tvar a rychlost šíření zůstávají po celou dobu téměř konstantní. Jak vápníkové vlny přecházejí z buňky do buňky, zůstává zčásti nezodpovězenou otázkou. Jisté je, že se šíří pouze buňkami, které jsou schopny vytvářet mezi sebou určitý typ spojů. A astrocyty jsou známé právě tím, že vytvářejí složité sítě buněk vzájemně propojených zvláštním typem kanálu (angl. gap-junction). Tento mezibuněčný spoj si buňky mohou podle potřeby otevřít, přivřít nebo zcela uzavřít. Zajímavé je, že jedním z faktorů, který na otevření komunikačního kanálu působí, je právě vápník. Mohli bychom si tedy představit, že zvýšení vápníkové koncentrace v jedné buňce otevře mezibuněčný spoj, kterým se vápník difuzí dostane do další buňky. Problém je v tom, že difuzní konstanta vápníku je příliš nízká. Kdyby se vápníkové vlny měly šířit rychlostí difuze vápníku, probíhal by celý proces daleko pomaleji. Další teorie je založena na předpokladu, že tím mezibuněčným kanálem neprochází vápník, ale IP3, jehož difuze intracelulárním prostorem je mnohonásobně rychlejší. Po difuzi do sousední buňky otevře IP3 vrátka jednoho z vápníkových rezervoárů, vápníková koncentrace se zvýší a tím stimuluje fosfolipázu C, která zvyšuje IP3 difundujícího do další buňky, a tam se celý proces opakuje. Vědci se potýkají s problémem, jak tuto teorii prokázat, neboť průběžné měření IP3 je tvrdým oříškem. (viz Vesmír 75, 137, 1996/3.)Mnoho experimentálních výsledků týkajících se vápníkových oscilací a vln bylo získáno na buněčných kulturách. Vlastnosti většiny buňek závisí totiž na tom, v jakém prostředí „vyrostly“. To platí pro všechny buňky, a pro gliové zvlášť. Proto byli zezačátku někteří badatelé na pochybách, zda tyto jevy neexistují jen v buňkách uměle pěstovaných, zda to není jen bláznivé chování nenormálně vyvinutých buněk. V posledních letech bylo proto věnováno značné úsilí k prokázání existence vápníkových oscilací a vln v co nejfyziologičtějších podmínkách a dnes již existuje dostatek experimentálních důkazů potvrzujících, že též in vivo vápníková signalizace formou oscilací a vln skutečně existuje.
K čemu je to všechno dobré
V minulé části jsme mluvili o neuro-gliové signalizaci a řekli jsme si, že jak nervové, tak gliové buňky jsou vybaveny arzenálem receptorů, neuropřenašečů i iontových kanálů, které jim umožňují spolu komunikovat. Dnešní názor na neuro-gliovou interakci je, že oba druhy buněk se dorozumívají způsobem, který se v lecčems podobá synaptickému přenosu (viz Vesmír 75, č. 3 a 4).Jedním z mechanizmů, kde se uplatňují principy známé ze synaptického přenosu v kombinaci s vápníkovou signalizací gliovými buňkami, je řízení energetického metabolizmu nervového systému. Neurony spotřebovávají značné množství energie, kterou si doplňují tím, že z krevního oběhu odebírají glukózu. Jenže neurony mohou být značně vzdáleny od místa, kudy procházejí kapiláry krevního oběhu. Mezi krevními kapilárami a neurony však existuje hustá síť, tvořená astrocyty s jejich dlouhými rameny. A zde právě je oblast, kde se může uplatnit vápníková signalizace. Hladový neuron uvolňuje glutamát, který, jak už víme, vyvolává v astrocytech vápníkové oscilace. Čím víc glutamátu neuron uvolní, tím vyšší bude frekvence vápníkových oscilací v nejbližším astrocytu, který tak pozná, „jak velký hlad“ má jeho neuron. Další fáze bude vyvolání vápníkové vlny. Ta přenese „volání o pomoc“ až k astrocytu, který je v kontaktu s krevní kapilárou. Zvýšená koncentrace vápníku v astrocytu spustí mechanizmus, který umožní odebrat z kapiláry potřebné množství glukózy a přetransportovat ji zpět až k hladovému neuronu. Ve skutečnosti se během tohoto transportu astrocyt postará o jakési první zpracování glukózy, a to tím, že ji přemění v laktát, aby ulehčil neuronům práci s „trávením“.
Část právě popsaného mechanizmu již byla experimentálně prokázána, zbytek je vysoce pravděpodobný. To, že nervová aktivita vyvolává vápníkové oscilace a vlny v okolních gliových buňkách, bylo prokázáno nezvratně. Je též jisté, že se tento mechanizmus spouští uvolňováním různých neuroaktivních látek z neuronů a nervových vláken. Přechod glukózy z krevních kapilár do astrocytů je považován za prokázaný stejně jako její přeměna v laktát před odevzdáním nervovým buňkám.
Tato hypotéza je jednou z mnoha, při kterých vápníková signalizace hraje důležitou roli. Je totiž třeba si uvědomit, že výlev většiny neuropřenašečů, které jsou roztroušeny i se svými receptory na nejrůznějších místech téměř všude po celém organizmu, je závislý na náhlém zvýšení hladiny intracelulárního vápníku. Lze tedy předpokládat, že vápníková oscilace nebo vlna vyvolá výlev neuroaktivní látky, která se v dané buňce nachází, a tím i aktivaci okolních receptorů. Tento mechanizmus se může uplatnit v nesčetných biologických pochodech. Je velice pravděpodobné, že vápníkové oscilace a vlny šířící se gliovými buňkami mohou ovlivňovat činnost synaptického přenosu, modulovat vedení akčních potenciálů nervovými vlákny, přenášet příkazy umožňující udržování stabilních podmínek v mimobuněčném prostoru kolem intenzivně pracujících neuronů a jistě ještě mnoho jiného. Je nesporné, že zatím teprve pomalu poodhrnujeme roušku, pod kterou je ukryta většina těch nejdůmyslnějších fyziologických a biologických funkcí a pochodů.
Poznámky
O autorovi
Petr Jirounek
Doporučujeme

Příliš otevřené dveře

Tranzistor na bázi bismutu 
