















Homologie
Představa homologie dvou struktur uvnitř jednoho organizmu je pěkně stará – na počátku byl nápad, že obratlovčí lebka vlastně odpovídá několika srostlým obratlům. Dosud se občas vedou spory, zda se touhle otázkou zabýval první Oken nebo Goethe, nakonec prý opravdu Oken. Příběhy o tom, jak je to vůbec napadlo, se liší jen nepatrně, ale přece: zatímco prostý přírodovědec Oken dostal tento nápad, když našel v lese kostru jelena, hofrát Goethe byl inspirován nálezem ovčí lebky na židovském hřbitově v Benátkách (dekadent!).
(dle Aleca Panchena)
Řada biologických pojmů přežívá z dob předevolučních a dodatečně jsou jim dodávány evoluční významy. Takový stařičký pojem se ovšem vzpírá, musí být různými způsoby kroucen, následkem čehož pak existuje mnoho alternativních, vzájemně se vylučujících koncepcí užívajících téhož termínu. Tato situace hraje nezanedbatelnou roli v utužování vědeckých vztahů: vědci vůbec nemusí poznat, že spolu vlastně nesouhlasí.
Všichni tušíme, co to je homologie – tušíme, že jsou jaksi znaky správné, homologické, na jejichž základě lze organizmy legitimně seskupovat, a znaky falešné, homoplastické (analogické, nehomologické). Takto definována má homologie cosi společného s původem, s historií, s fylogenezí, a týká se dvou a více druhů. Ale taky můžete číst, že kupříkladu kladélko samic jednoho druhu hmyzu je (částečně) homologické penisu samců téhož druhu; nebo že hmyzí tykadlo buď je, anebo není homologické hmyzí noze. Je zde jakási homologie uvnitř druhu nebo i uvnitř jedince. Tato homologie jako by spíše odkazovala na sdílené genetické a ontogenetické procesy.
Jsou tu tedy dvě homologie, jedné se říká historická nebo fylogenetická (anebo ortologie), druhé biologická nebo iterativní (anebo paralogie), dále dělená na seriální, sexuální, ontogenetickou apod. Zdálo by se, že ty homologie nemají nic společného. Ale přece – dvě jednotky na jednom těle jsou si biologicky homologické, protože
- buď vznikly duplikací původně jedné jednotky u předka (např. geny pro α- a β-hemoglobin),
- anebo pocházejí ze dvou nebo více jednotek na těle předka, které ovšem byly plus minus stejné a teprve později se diferencovaly (křídlo a noha slepice). Takže odkaz k předkovi, k historii, je ve hře vždy; nicméně v tomto textu bude řeč pouze o homologiích historických. Nadále budu mluvit o homologických strukturách, protože o těch se nejlíp mluví, ale ve skutečnosti jde o obecně paradigmatický pojem, který lze (a který je třeba) aplikovat i na vývojové dráhy, vzorce chování, ekologické interakce, zkrátka na cokoliv, co má nějakou evoluci.
Pojem „homologie“ se svádí na Richarda Owena (1843), ale ono je to samozřejmě složitější. To, čím se Owen proslavil, srovnávání lidských, ptačích a krokodýlích kostí a nalézání homologických vztahů mezi nimi, je mnohem staršího data. Stál-li Owen oběma nohama v době evoluční (přičemž ovšem s Darwinem si šli tak strašlivě na nervy, že dodnes jsou v londýnském přírodovědném muzeu sice vytesáni z bílého mramoru oba, ale umístěni daleko od sebe, aby se neservali), jakýsi pan Belon publikoval srovnání lidské a ptačí kostry už roku 1555 a divil bych se, kdyby on byl první, koho to napadlo, ono je to nasnadě. Co tedy praví Owen? Překvapivě skoro nic, pouze v glosáři své práce „Lectures on Comparative Anatomy and Physiology of the Invertebrate Animals, Delivered at the Royal College of Surgeons in 1843“ definuje homologii dosti mlhavě jako „stejný orgán u různých živočichů jakkoliv rozdílný ve formě a funkci“ („the same organ in different animals under every variety of form and function“), zatímco analogii (tj. ne-homologii), jako „část nebo orgán jednoho živočicha, který má stejnou funkci jako jiná část nebo orgán jiného živočicha“ („a part or organ in one animal which has the same function as another part or organ in a different animal“). Takže jenom vědět, co znamená „the same“, „different“, „another“ a „every variety“. Posledních 153 let se vedou spory jenom o tato čtyři slova, to ostatní už máme pod střechou. Tak kupříkladu slavný paleontolog L. Van Valen praví, že homologie je „shoda způsobená kontinuitou informace“. A L. Rothová akceptuje tuto definici s nelíčeným nadšením a volá: „tuto definici může užít libovolná myšlenková škola prostě tak, že určí relevantní druh informace“. No právě. Každý pojem si můžeme definovat buď tak, abychom o něm mohli vést učené řeči (tedy co nejobecněji), nebo tak, abychom ho mohli používat (tedy co nejoperativněji, obvykle dosti úzce). Van Valenova definice očividně patří k těm, které vznikají proto, aby se každý, kdo na nějaké ne zcela vzdálené téma někdy něco utrousil, dočkal satisfakce, že na jeho slova došlo.
Historické homologie nehledáme jenom proto, že z toho máme plezír (viz níže), ale hlavně proto, abychom je, jako důsledky historické kontingence (Neubauer – Vesmír 74, 646, 1995/11), mohli vzít a zahodit, protože nám, jako důsledky historické kontingence, o obecných zákonitostech tohoto světa neříkají nic. Znak, který svou historií kopíruje fylogenezi, nám poskytuje informaci o fylogenezi, ale stěží něco víc. Nejprve je třeba odfiltrovat všechny fylogenetické náhody, všechno, co je, ale být by nemuselo, abychom mohli zkoumat ten zbytek; jinak nemá cenu hledat nějaké funkční nebo genetické (od „geneze“, ne „genetika“) vysvětlení koexistence dvou znaků jednoho druhu (třeba mezi žitím ve vodě a dýcháním žábrami).
Dám jednoduchý příklad: Je na světě jeden jediný stát, jehož hlavní město se jmenuje Praha, a jeden jediný stát, který má ve státním znaku stříbrného dvouocasého lva; a co byste neřekli, je to tentýž stát. Jaká je tedy funkční korelace mezi slovem „Praha“ a stříbrným dvouocasým lvem? Žádná, že? – Pojďme dál: je na světě nějakých 40 000 druhů, které mají snovací bradavky na zadečku, a nějakých 40 000 druhů, které užívají druhý pár končetin (pedipalpy) k přenosu spermií; a co byste neřekli, je to těch samých 40 000 druhů (tzv. pavouci). Jaká je tedy funkční korelace mezi snovacími bradavkami a přenosem spermií pomocí pedipalpů? Zase žádná ... nebo aspoň nemůžeme říct, jaká a zda vůbec nějaká. Všichni pavouci totiž pocházejí od společného předka, který už obě vlastnosti měl. 40 000 pavouků tedy nepředstavuje 40 000 nezávislých opakování, 40 000 nezávislých experimentů provedených Matkou Přírodou, které bychom mohli statisticky zkoumat (a divit se stoprocentnímu výsledku), ale prostě jeden jediný, unikátní, jedinkrát se přihodivší experiment, Praotec Pavouk, ve 40 000 kopií. Jednou jedinkrát se zkrátka snovací bradavky a pářicí pedipalpy objevily najednou u jednoho druhu – co s tím chcete dělat jiného, než to prostě konstatovat? A jaká je funkční nebo genetická souvislost mezi srpovitými chelicerami a produkcí jedu v nich, když tyto znaky zase tvoří unikátní evoluční novinky charakterizující všechny potomky jednoho jediného Praotce Pavouka? Nějaká souvislost tam asi bude – ale dokažte to; prostá kontingence představuje nulovou hypotézu a je třeba ji vyvrátit.
To, že existuje korelace mezi přítomností dvou znaků (třeba žití ve vodě & dýchání žábrami, nápadné zbarvení & nepoživatelnost, nebo velká varlata & vysoká promiskuita), má nějakou výpovědní hodnotu pouze tehdy, věříme-li, že druhy, které oba tyto znaky nesou, netvoří jednu monofyletickou skupinu. Asi řeknete, že asociace barevnosti a nechutnosti něco znamená, protože skupina barevných a nechutných zvířat monofyletická není. Ano, a jak to víte? No přece tak, že uznávaná fylogenetická hypotéza vypadá jinak. Nejdřív fylogeneze, potom identifikace homologií a jejich neutralizace – a potom se uvidí.
Hledáme-li operativní kritéria, jak poznat homologickou podobnost od nehomologické, jsou nám nabídnuty kupř. poloha, ontogeneze, speciální podobnost. Pokaždé jde o apriorní kritéria, která by měla být použitelná, když se díváme tzv. poučenýma očima na organizmy, jejichž fylogenetické vztahy ještě znát nemusíme. Všechna tato kritéria mají cosi společného – kromě případů, které jednoznačně akceptují (noha mouchy je homologická noze brouka), a případů, které jednoznačně odmítnou (noha mouchy není homologická noze prasete), zanechávají znepokojivě širokou šedou zónu, kde kupodivu různé autority vidí svýma poučenýma očima velmi různé věci (je noha mouchy homologická parapodiu mnohoštětinatce?). Kromě toho se nemohu zbavit dojmu, že jakmile se věda začne obracet na moudré rabíny, kteří jedině mohou rozhodnout tam, kde my tápeme, přestává být vědou.
Homologie pro sdílení polohy praví: dva, třeba i nepodobné, orgány vyrůstající ze stejného místa jsou si homologické. Kde má moucha kyvadélka, mají jiní hmyzové zadní křídla (historická homologie), a kde má moucha na zadohrudi kyvadélka, tam má na středohrudi křídla (biologická homologie), čili kyvadélka jsou homologická křídlům. Jenže – přední končetiny mloka, ještěrky a člověka jsou si (očividně) homologické, ale bohužel rostou na různých místech. Každý, kdo kdy tepelně upravoval rybu, si všiml, že se mu rozpadá napříč, a to na mnoha predeterminovaných místech: hezký – takový empirický – důkaz faktu, že obratlovci jsou segmentovaní jako žížaly, snad jen že u vzrostlého člověka už to tak pěkně nevypadá, s výjimkou páteře, míšních nervů a příslušných svalů. Nuže, z kterých tělních segmentů vyrůstají přední nohy obratlovců? Z 2. – 5. segmentu mloka, 6. – 9. segmentu ještěrky a 13. – 18. segmentu člověka. Takže buď homologické končetiny vyrůstají z nehomologických míst (což by nemělo být), anebo nejsou homologické.
Homologie pro sdílení speciální podobnosti je něco podobného: ruka člověka a křídlo netopýra jsou si globálně dosti nepodobné (zjevně proto, že netopýr si v evoluci opomenul uvolnit přední končetiny pro práci), ale detailní analýza (třeba počtu a uspořádání kostí) ukazuje, že rozdíly jsou spíše kvantitativní. Ale – kde se zastavíme? Je dostatečnou speciální podobností hmyzí nohy a tykadla jejich segmentace? (Ano.) Vadí, že hmyzí kusadlo segmentované není, a noha je? (Ne. Kusadlo ztratilo distální segmenty.) Je dostatečnou speciální podobností hmyzí nohy a křídla to, že to obojí ční z těla? (Ne. I když ... data molekulární biologie budí jisté rozpaky.)
A zbývá tedy stejná (plus minus stejná) ontogeneze jako kritérium homologie.
Co už ale víme tak tvrdě, jak lze v postmoderním světě vůbec něco vědět – homologické struktury nesporně vznikají velmi různě. Vůbec to nemusí vypadat tak, jak si lidé obvykle myslí, že zpočátku podobné dráhy se postupně rozcházejí, divergují, a vytvářejí rozdílné (ale homologické) struktury. Naopak – často jsou právě raná stadia ontogeneze (rýhování, gastrulace, celomogeneze) nesmírně rozdílná, dokud se nesejdou (ne-v-konvergují) ve fázi invariantní. A občas má evoluce (koho čeho:) ontogeneze tvar přesýpacích hodin – raná embrya a dospělci jsou velmi variabilní, a někde uprostřed je stadium (obvykle pořád ještě embryonální) o vysokém stupni invariance (tzv. fylotypické stadium). A vraťme se k homologiím: Co když to znamená, že jen některá stadia mají předepsáno být evolučně stabilní, zatímco všechna ostatní se mění, jak se to zrovna hodí, anebo dokonce (obvykle?) náhodou. Jak potom užívat společnou ontogenetickou dráhu dvou orgánů jako důkaz jejich homologie? Myslím si zkrátka, že homologický výlet do Vídně může být uskutečňován nehomologickými prostředky, jako jsou hůl a dobrý boty, kůň, nebo zeppelin.Ještě jednou se vraťme k již citovanému příkladu vzniku oční čočky mloků (Vesmír 74, 145, 1995/3). Čočka embryonálně vzniká z ektodermu, ale regeneruje z mezodermální duhovky. Dovoluji si připomenout, že vzniká-li čočka jednou z ektodermu a jindy z mezodermu, není to jakási drobná odchylka od úplné totožnosti. Můžete namítnout, že prostě embryogeneze a regenerace jsou dvě různé věci. No jistě, ale tady jde přece o ten vývojový potenciál. Jsou-li dvě nepříbuzné tkáně schopny vyrobit, byť v různou dobu, tutéž funkční strukturu, proč by se tato schopnost nemohla fylogeneticky rozštěpit. Co když v rámci obratlovců vznikne (nebo vznikl) druh, jehož oční čočka bude se svým vznikem čekat na diferenciaci duhovky a vznikne až z ní? Bude oční čočka tohoto druhu (a jeho potomků, tedy čeledi, třídy, kmene ...) homologická čočce ostatních obratlovčích skupin? A když řeknete, že ano, řekněte ještě, jak tu homologii poznáte, až ty dvě různě vyráběné oči dostanete k analýze a nebudete a priori vědět, jak to vznikalo? Což a priori vědět vskutku nebudete. 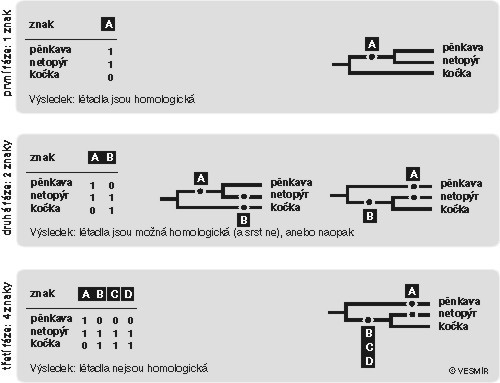
Můžeme tedy sumarizovat: Homologie skrz sdílenou vývojovou dráhu někdy platí, někdy ne a my nevíme, kdy ne; homologie skrz polohu a homologie skrz detailní morfologii jsou na tom ještě hůř. A sumarizujme ještě dál: Neexistuje žádná metoda, jak spolehlivě a priori rozpoznat homologii od nehomologie; existují pravda různé pocity, ale ty nemusejí být sdíleny více pracovníky. Pohledem na organizmus, ani pohledem na dva organizmy nemůžeme homologii rozpoznat, ani kdybychom je kvůli tomu prohlíželi elektronovým mikroskopem, zkoumali od nejranějšího embrya, strkali do nukleárního magnetického rezonátoru (ať už takový inštrument dělá cokoliv), či rozmělnili až na molekuly. Musíme s nimi udělat něco úplně jiného.
Metoda aposteriorní říká, že co je homologie a co není, nemůžeme poznat, a proto s každou podobností budeme předběžně pracovat, jako by to homologie byla. Čili máme-li pěnkavu, netopýra a kočku, všechny podobnosti mezi jakoukoliv dvojicí druhů považujeme za potenciálně homologické; to neznamená, že tomu máme věřit, ale že je to nezbytný metodologický přístup (obr. obrázek). Jsou křídlo pěnkaví a křídlo netopýří homologická, a je tedy třeba pěnkavu a netopýra zařadit do jedné skupiny a kočku nechat zvlášť? Ano – pokud s tím jiné znaky nejsou v rozporu. A to jsou: kočka a netopýr mají více společných znaků (a proto jsou to savci) než pěnkava a netopýr (a proto to nejsou létavci). Jako obvykle tedy vítězí většina – mléčné žlázy, srst a bezjaderné erytrocyty přehlasovaly „přední okončetiny přeměněné v létadla“, jak pěkně praví Brehm. Protože teď a tady se hrálo o poctu být uznán homologickým, mléčné žlázy, srst a bezjaderné erytrocyty jsou homologické, ale létadla ne. (Dodávám – nejde o sdílení znaků, ale pouze odvozených znakových stavů, tzv. synapomorfií. A ještě dodávám – ptačí a netopýří křídlo jsou si homologické jako končetiny, ale nehomologické jako létadla.)Čili – homologické jsou ty znaky, které se v evoluci mění spolu (věříme-li na evoluci), nebo ty znaky, které společně definují skupinu druhů jako výsek obecné hierarchie všeho živého (vidíme-li kolem sebe objektivně existující a popisovatelnou hierarchii, ale už necítíme potřebu spekulovat o jejím vzniku). Homologické jsou ty znaky, které vytvořily koalici, a to ne jen tak ledajakou, ale koalici největší, koalici převažující. Homologie je výsledek fylogenetické analýzy (obr.
Jsou ovšem lidé, kteří říkají zhruba toto: Homologické znaky musejí být rozlišeny dříve, než se do fylogenetické analýzy pustíme, protože jen analýza homologických znaků nám dá tu pravou fylogenezi. Některé znaky zkrátka nesou fylogenetickou informaci, jiné jenom šum, a to je potřeba odlišit předem. Nu. Za prvé – žádná „pravá fylogeneze“ neexistuje, nějaká sice byla, ale není jiný nástroj, jak ji identifikovat, než analýza, z které nám prostě něco vyleze. Ptát se, zda je výsledná fylogenetická hypotéza „ta pravá“ nemá smysl, pokud se tím vlastně neptáme, zda jsme ji vytvořili správně. I fylogenetiku lze – jak vidět z mnohých případů – dělat špatně; výsledek správně provedené analýzy je pak ... inu, prostě správný, není-li dokázán opak.
Za druhé, kritéria, jak rozlišit znaky a priori dobré od a priori špatných sice existují, ale je s nimi svízel, jak ukázáno výše. Nebudou-li zastánci apriorní homologie schopni navrhnout nic lepšího než speciální podobnost, polohu a ontogenezi, nebo nebudou-li schopni navrhnout metodu, jak poznat, kdy tato kritéria platí a kdy ne, není se s nimi jaksi o čem bavit. Představa, že existuje tzv. zdravý rozum, že přece všichni víme, jak to na tom světě chodí, že netřeba trvat na nějakých (zde metodologických) formalitách, že všichni normálně vnímající lidé přece vidí to samé, a mohou se tedy, když je dobrá vůle, takňák dohodnout, se nazývá naivní realizmus a široce rozšířena je zvláště mezi řezníky a ženami v domácnosti.
A za třetí, dobrá, dobrá, nebude hned tak zle – předáme matici znaků nějakému aprioristovi, aby z ní vyhodil všechno, o čem (si myslí, že) ví, že to nejsou homologie. Zbude-li v matici po této operaci větší počet znaků (dejme tomu více než 20), dám ruku do ohně, že nastane jeden ze dvou případů:
- Analýza opět některé z nich odhalí jako „homologické“ (tedy kongruentní s výslednou fylogenetickou hypotézou) a jiné jako „nehomologické“, čili zase nějaký konflikt mezi těmi teď už opravdu pravými homologiemi, což by mělo aprioristovi postačit, aby si uvědomil, že jeho kritéria prostě neplatí.
- Anebo všechny znaky vyjdou jako „homologické“, což těžko vysvětlit jinak, než že apriorista hodnotil homologii-nehomologii srovnáváním s předem přijatou fylogenetickou hypotézou, že tedy nepracoval apriorně, nýbrž aposteriorně („sdílená nepřítomnost zubů u mravenečníků a velryb je irelevantní, protože víme, že mravenečníci i velryby vznikli nezávisle na sobě z ozubených předků“). Že tedy dělal to, co nám předtím vyčítal. Snad jen že to dělal intuitivně, nahradiv striktní metodu svým dobře vyvinutým poučeným zrakem, což se v jistých kruzích považuje za přednost.
Dejme tomu, že máme dva druhy myší, jedna má délku těla 8 cm a délku ocasu 8 cm, druhá má délku těla 8 cm a délku ocasu 4 cm. Je zřejmé, že se čímsi liší, patrně nějakými znaky. Kolika? Znak A („délka těla“) nabývá jediného stavu (8 cm), a můžeme jej tedy opominout. Znak B („délka ocasu“) má dva stavy, 8 cm a 4 cm; a pak je tu taky znak C („poměr délky těla k délce ocasu“) s dvěma stavy, 1:1 a 2:1. Tedy tři znaky, že. Všichni snad tuší, že ne, že znaky B a C jsou totéž. Ale co když máme dva druhy něčeho, jeden s malou hlavou a jeden s velkou. Je to jeden znak, nebo tolik znaků, kolik jich jenom dokážeme změřit, nebo pouze některé pěkné rozměry uznáme za znaky? Nedávno vyšla morfometrická práce o tvaru včelích křídel v závislosti na velikosti těla. Skoro všechny rozměry, které taxonomové na křídelní žilnatině naměřili (dejme tomu 20 znaků) a které mimo jiné užívali pro klasifikaci včel, jsou závislé na velikosti těla. Je to tedy 20 + 1 znak (což si taxonomové až dosud mysleli), nebo jediný znak, „velikost těla a její morfometrické konsekvence“? To samozřejmě nevím: musíme se podívat do genů a do ontogenetických trajektorií, což nikdo dosud neudělal, takže nevíme, co je znak.
- Nemá cenu hovořit o homologii v tom širokém smyslu, jak se dosud děje. Je asi rozumné akceptovat původně molekulární pojmy ortologie a paralogie pro různé druhy „homologií“, nebo užívat pojem „homologie“ tam, kde ho všichni jaksi cítíme jako nejvhodnější: „homologie“ = historická homologie = ortologie.
- Nelze poznat homologii předem, homologie nám prostě vyjde, anebo nevyjde. Kritériem evoluční správnosti znaku, jeho homologičnosti, je kongruence jeho taxonomické distribuce s taxonomickou distribucí jiných znaků.
- Ontogeneze a genetika nám pomohou rozpoznat, co je znak, ale ne co je homologie. Protože znak je zkrátka nějaký takový tento, o němž předpokládáme, že prodělává evoluci víceméně nezávislou na podobných takových tento, a aby to mohl činit, je víceméně nezávisle geneticky řízen a ontogeneticky vytvářen; kdežto homologie je takový znak, který svou evolucí sleduje fylogenezi organizmů.
- Nic, ale vůbec nic, se nerozumí samo sebou. Se vším se musí dělat věda.
Poznámky
Znak je primitivní/odvozený proto, že to odpovídá jeho distribuci na fylogenetickém stromu. Jakákoliv alternativní metoda je buď špatná („jednoduché je primitivní“, „obecně rozšířené je primitivní“, „paleontologicky staré je primitivní“), anebo sporná a podezřelá („ontogeneticky rané je primitivní“). Neexistuje žádná „metoda paleontologická“ a existence „metody ontogenetické“ je pochybná (Horáček – Vesmír 74, 453, 1995/8); paleontologie a vývojová biologie pouze dodávají údaje, které by prostému recentnímu a adultnímu morfologovi chyběly, ale tyto údaje samy o sobě nenesou žádnou informaci o fylogenezi, dokud nejsou analyzovány úplně stejně jako údaje morfologické (tj. „metodou srovnávací“). Fosilie, pravda, představují „evoluci zamrzlou v horninách“, stejně jako žijící organizmy představují „evoluci zamrzlou v bílkovinách“; o to jsou fosilie jako zdroje informací horší, nikoliv lepší. Zásadní přednost fosilií proti recentním organizmům by měla být v tom, že obsahují údaje o samotném plynutí času – ale abychom nezačali hovořit o změnách, které proběhly od prvohorních ramenonožců k druhohorním mlžům, musíme vědět, že ramenonožci a mlži jsou dvě různé skupiny, musíme mít fylogenetickou hypotézu, a kde jsme ji vzali? Inu, „metodou srovnávací“, ta se totiž nemusí omezovat na jediný časový řez, jak naznačuje Horáček, protože této metodě do nějakého času nic není. Že z takto pojatého světa mizí čas (nikoliv čas geologického datování, ale čas, v němž se dějí příběhy), je pravda a já myslím, že je to tak dobře. Jinak jsme totiž odsouzeni k vypravování histórií, což některým vadí.
O autorovi
Jan Zrzavý

Doporučujeme

Příliš otevřené dveře

Tranzistor na bázi bismutu 
