














Kdo jsme, odkud přicházíme a kam směřujeme
To jsou otázky, které si klademe odnepaměti. V umělecké rovině se s nimi pokusil vyrovnat například Paul Gauguin ve svém slavném obrazu, jehož název mě inspiroval pro titul článku.
Kdo jsme
Asi každý, ať už vychází z jakýchkoliv předpokladů, připustí nápadnou podobnost lidí a vyšších primátů, zejména pak naši podobnost se šimpanzi. Nejde jen o celkovou tělesnou stavbu, ale také o různé projevy inteligence. Šimpanzi vyrábějí nástroje, což je celkem běžné i u řady jiných zvířat, navíc však vytvářejí i kulturní tradice a vedou dokonce války. U člověka je ovšem všechno řádově umocněno. Na druhou stranu je mezi námi a šimpanzi přeci jen pořádná mezera. Definování rozdílů, které ji působí, může mnohé napovědět o podstatě člověka jako biologického druhu i sociální bytosti.
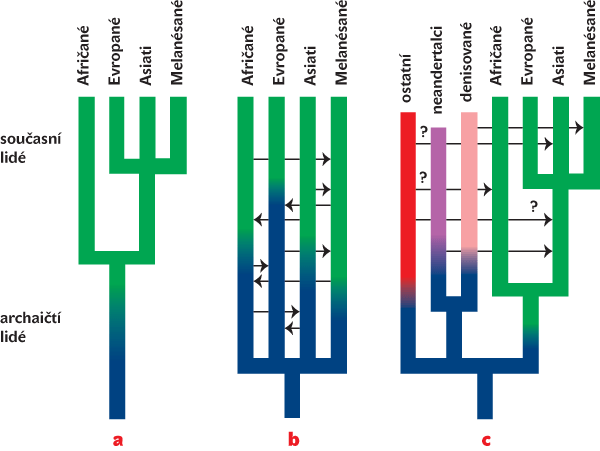
 Jedinečnou příležitost k analýze na kvalitativně nové úrovni přineslo přečtení úplné genetické informace – všech písmen v knize života – tedy lidského i šimpanzího genomu.1) Nedávno se navíc podařilo s pomocí nejmodernějších sekvenačních postupů2) určit úplnou sekvenci – pořadí nukleotidů – mitochondriální DNA (mtDNA) a posléze i celého jaderného genomu neandertálce. Ukázalo se, že šimpanzí a lidský genom se velmi podobají (z 98,5 %), což je významně větší shoda než třeba u příbuzných druhů hlodavců – myši a potkana. Dalším překvapením bylo, že lidský genom obsahuje poměrně málo genů kódujících přímo proteiny. Očekávalo se alespoň 100 000. Skutečných 21 000 genů je v živočišné říši celkem průměrný počet. Další analýzou získaných dat bylo v lidském genomu brzy nalezeno poměrně velké množství genů s významnými strukturálními změnami ve srovnání se šimpanzem. Často jde o transkripční faktory, tedy o regulátory exprese3) velkého množství dalších genů.
Jedinečnou příležitost k analýze na kvalitativně nové úrovni přineslo přečtení úplné genetické informace – všech písmen v knize života – tedy lidského i šimpanzího genomu.1) Nedávno se navíc podařilo s pomocí nejmodernějších sekvenačních postupů2) určit úplnou sekvenci – pořadí nukleotidů – mitochondriální DNA (mtDNA) a posléze i celého jaderného genomu neandertálce. Ukázalo se, že šimpanzí a lidský genom se velmi podobají (z 98,5 %), což je významně větší shoda než třeba u příbuzných druhů hlodavců – myši a potkana. Dalším překvapením bylo, že lidský genom obsahuje poměrně málo genů kódujících přímo proteiny. Očekávalo se alespoň 100 000. Skutečných 21 000 genů je v živočišné říši celkem průměrný počet. Další analýzou získaných dat bylo v lidském genomu brzy nalezeno poměrně velké množství genů s významnými strukturálními změnami ve srovnání se šimpanzem. Často jde o transkripční faktory, tedy o regulátory exprese3) velkého množství dalších genů.
Pomohla nám řeč?
 I přes pozoruhodné šimpanzí výkony v komunikačních schopnostech, včetně zvládnutí velkého množství symbolů znakové řeči, je lidská mluva nesrovnatelně účinnější a představuje zcela jistě evoluční novinku. Hledání genů určujících tuto vlastnost je tedy jednou z možných strategií pro objasnění otázky, kterou jsme si položili v úvodu. Podstata neuromolekulárních drah, které jsou základem jazyka, není známa, ale velký informační význam může mít poziční klonování4) lidských neurologických vývojových vad. Pokud mutace daného genu odpovídá za poruchu určité neurální funkce, je pravděpodobné, že sekvenční variace v tomto genu mohou ovlivňovat příslušnou vývojovou cestu. Srovnání vnitrodruhové a mezidruhové variability (šimpanze a člověka) pak může posloužit k vyvození závěrů, zda příslušný gen hraje významnou roli v evoluci člověka.
I přes pozoruhodné šimpanzí výkony v komunikačních schopnostech, včetně zvládnutí velkého množství symbolů znakové řeči, je lidská mluva nesrovnatelně účinnější a představuje zcela jistě evoluční novinku. Hledání genů určujících tuto vlastnost je tedy jednou z možných strategií pro objasnění otázky, kterou jsme si položili v úvodu. Podstata neuromolekulárních drah, které jsou základem jazyka, není známa, ale velký informační význam může mít poziční klonování4) lidských neurologických vývojových vad. Pokud mutace daného genu odpovídá za poruchu určité neurální funkce, je pravděpodobné, že sekvenční variace v tomto genu mohou ovlivňovat příslušnou vývojovou cestu. Srovnání vnitrodruhové a mezidruhové variability (šimpanze a člověka) pak může posloužit k vyvození závěrů, zda příslušný gen hraje významnou roli v evoluci člověka.
Například studium primární mikrocefalie (vývojové poruchy vedoucí k redukci objemu mozku) naznačuje mechanismus, který v evoluci člověka vedl k expanzi kortexu, vnější vrstvy savčích mozků, odpovědné za jeho vyšší kognitivní funkce. Mezi geny, které ovlivňují velikost mozku, jsou významné regulátory buněčné proliferace a buněčné smrti MCPH1 (kódující mikrocefalin) a ASPM (abnormal- -spindle-like, microcephaly associated), který je důležitý pro formování vřeténka při dělení neuroblastů. Tyto geny vykazují známky pozitivní selekce v evoluci primátů.5) Mikrocefalici ovšem mluví celkem normálně a jsou jen mírně mentálně retardovaní.
 Přímější pohled na evoluci jazyka poskytují studie vývojových poruch bezprostředně zasahujících řečové schopnosti. Nejvýznamnější se jeví gen pro transkripční faktor FOXP2 (forkhead box P2). Mutace v DNA vazebné doméně FOXP2 způsobuje poruchu řeči, při které je ovlivněna schopnost jemných pohybů úst a hrtanu. Dvě klíčové záměny aminokyselin FOXP2 proteinu mohou být důvodem neschopnosti artikulace u šimpanze.6) Z jiných významných genů jmenujme ještě MYH16, který je nutný pro vývoj žvýkacích svalů. Mutace u člověka vedla k zeslabení čelistí, které mohlo umožnit růst lidské lebky. Další nápadné odlišnosti jsou v genech ovlivňujících čich a sluch, některých genech souvisejících s imunitní odpovědí a genech spojených s reprodukcí.
Přímější pohled na evoluci jazyka poskytují studie vývojových poruch bezprostředně zasahujících řečové schopnosti. Nejvýznamnější se jeví gen pro transkripční faktor FOXP2 (forkhead box P2). Mutace v DNA vazebné doméně FOXP2 způsobuje poruchu řeči, při které je ovlivněna schopnost jemných pohybů úst a hrtanu. Dvě klíčové záměny aminokyselin FOXP2 proteinu mohou být důvodem neschopnosti artikulace u šimpanze.6) Z jiných významných genů jmenujme ještě MYH16, který je nutný pro vývoj žvýkacích svalů. Mutace u člověka vedla k zeslabení čelistí, které mohlo umožnit růst lidské lebky. Další nápadné odlišnosti jsou v genech ovlivňujících čich a sluch, některých genech souvisejících s imunitní odpovědí a genech spojených s reprodukcí.
Velice významné je porovnání projevů genů v různých tkáních. Zatímco třeba v játrech a v krvi nebyly zaznamenány žádné rozdíly, mozek šimpanze a člověka je na první pohled významně odlišný. V lidském mozku jako by se využívání mnoha genů zrychlilo. Navíc se v komplexu mitochondriálních genů (Complex IV – COX) odpovědných za transport elektronů nahromadilo v lidské linii výjimečně mnoho změn. Oba tyto nálezy mají jistě souvislost s energetickým metabolismem v mozku. Je ale otázkou, zda uvedené změny primárně umožnily růst mozku, nebo jsou až sekundárním důsledkem zvětšení mozku a jeho extrémní energetické náročnosti.
Potřebné haraburdí
 Možná tím nejdůležitějším hnacím motorem evoluce, alespoň v některých případech, jsou oblasti mimo vlastní geny. Zprvu byly považovány za balastní DNA (zvanou v anglické literatuře junk-DNA, doslova odpad, haraburdí). Vzhledem k tomu, že geny kódující proteiny tvoří jen asi 2 % sekvencí, bylo by podivné, kdyby zbylých 98 % bylo jen přítěží. Nabízí se porovnat úroveň poznání struktury a fungování genomu s okolním vesmírem. Najednou jsme konfrontováni se skutečností, že většina genomu sestává ze sekvencí, jejichž funkce je nejasná (temná), ale vypadá to, že mají rozhodující úlohu v evoluci. Také vesmír je, zdá se, tvořen převážně temnou hmotou a energií neznámé povahy, které rozhodují o jeho osudu.
Možná tím nejdůležitějším hnacím motorem evoluce, alespoň v některých případech, jsou oblasti mimo vlastní geny. Zprvu byly považovány za balastní DNA (zvanou v anglické literatuře junk-DNA, doslova odpad, haraburdí). Vzhledem k tomu, že geny kódující proteiny tvoří jen asi 2 % sekvencí, bylo by podivné, kdyby zbylých 98 % bylo jen přítěží. Nabízí se porovnat úroveň poznání struktury a fungování genomu s okolním vesmírem. Najednou jsme konfrontováni se skutečností, že většina genomu sestává ze sekvencí, jejichž funkce je nejasná (temná), ale vypadá to, že mají rozhodující úlohu v evoluci. Také vesmír je, zdá se, tvořen převážně temnou hmotou a energií neznámé povahy, které rozhodují o jeho osudu.
Ve struktuře genomu přece jen již začínáme něčemu rozumět. Samostatnou kapitolou by byla úloha různých typů krátkých regulačních RNA, jejichž seznam neustále roste a je zřejmé, že daleko největší část genomu je přepisována do nekódující RNA. Velmi zajímavá je také úloha retroelementů,7) které jsou vlastně genovými parazity, nicméně se zřejmě podílejí významnou měrou na utváření nejen lidského genomu. Nedávné studie ukázaly, že zvětšení našeho genomu urychlily právě retrotranspoziční aktivity. V genetické linii vedoucí k dnešnímu člověku vzrostl za posledních 50 milionů let genom o 30 Mb (megabází) ve srovnání se šimpanzem a o 550 Mb v porovnání s lemurem. Nejméně 90 % je způsobeno novými inzercemi retroelementů. Typickými představiteli jsou u člověka L1 a Alu, které představují asi 27 % lidského genomu. Nápadné je hlavně zvýšení aktivity Alu v posledních 25 milionech let. Tyto sekvence mohou představovat významné mutageny na strukturální i genové úrovni a jejich příspěvek ke změnám fenotypu může být větší, než se dříve myslelo. Také je zajímavé, že aktivita sekvencí Alu je velmi malá v nedávno publikovaném genomu orangutana. I jiné charakteristiky jeho genomu svědčí o relativně malé evoluční dynamice.
V poslední době byly přineseny další důkazy o specifickém zrychlení evoluce některých oblastí lidského genomu nazývaných HAR (Human Accelerated Regions). Celkem bylo nalezeno 49 úseků HAR. Z toho jsou pouze dvě klasické kódující oblasti. Z nekódujících sekvencí však celých 25 % sousedí s geny, které se podílejí na embryonálním vývoji mozku. Zrychlení evoluce ilustruje oblast nazvaná HAR-1 o velikosti 118 bp (párů bází). V této sekvenci byly nalezeny dvě odlišné báze mezi kuřetem a šimpanzem, které odděluje asi 310 milionů let separátního vývoje. Jenže šimpanz a člověk se odlišil v 18 bázích, a to za pouhých 6 milionů let. Oblast HAR-1 nemá kódující proteinovou kapacitu, ale transkript se pravděpodobně může skládat do stabilní struktury RNA. HAR-1 je aktivní v průběhu 7.–19. týdne těhotenství v Cajal-Retziusových neuronech, které jsou odpovědné za vytváření šestivrstevné mozkové kůry typické pro člověka. Celkově se tedy zdá, že největší rozdíly mezi příbuznými druhy budou spočívat, alespoň v některých případech, spíše v orchestraci celého souboru genů než v jednotlivých rozdílech uvnitř genů či v jejich celkovém počtu.
Za kritické přečtení textu děkuji RNDr. Jiřímu Hejnarovi.
Pokračování ve Vesmíru 2011/9.
Poznámky
1) První nástin kompletní lidské sekvence DNA byl publikován právě před 10 lety, patřičná pozornost tomu byla věnována v posledních číslech časopisů Science a Nature.
2) Mezi nové metody „čtení“ DNA patří 454 sekvenování nabízené firmou 454 Life Sciences. Metoda je založena na pyrosekvenaci, při níž je začlenění určitého nukleotidu („písmena“) do vznikajícího řetězce DNA doprovázeno světelným signálem v důsledku uvolnění pyrofosfátu, kdy následně vzniká ATP, který využije enzym luciferáza pro oxidaci luciferinu. Příprava na samotnou sekvenaci zahrnuje několik důmyslných kroků. Na paramagnetické kuličky potažené streptavidinem jsou navázány specifické biotinylované oligonukleotidy. Kuličky mají průměr 1 μm a jejich magnetických vlastností se využívá pro snadnou separaci. V olejové mikroemulzi jsou kuličky smíchány s fragmenty studované DNA. V kapénkách, kde je přítomna kulička a jeden fragment DNA, proběhne pomocí PCR (polymerázové řetězové reakce, viz Vesmír 77, 444, 1998/8) amplifikace iniciovaná oligonukleotidem navázaným v předchozích krocích. Po odstranění emulze a denaturaci DNA, jsou kuličky nesoucí jednovláknovou (klonální) DNA přeneseny centrifugací do pikolitrových jamek na mikrodestičce sestavené z optických vláken. Do každé jamky se vejde právě jedna kulička. Poté jsou přidány ještě menší kuličky nesoucí imobilizované enzymy potřebné pro pyrosekvenaci. Destička má hustotu jamek 480 na mm2 a obsahuje asi 1,6 milionů jamek. V sekvenačním přístroji je umístěna do průtokové komůrky se sekvenačními reagenty. Detekční systém je uzpůsoben k zachycení emitovaných fotonů z každé individuální jamky. Paralelně probíhá obrovské množství sekvenačních procesů najednou a výsledek vyhodnocuje výkonný počítač. Pro srovnání: první přečtení genomu člověka před deseti lety trvalo konsorciu HGP a společnosti Celera Genomics několik let a stálo asi tři miliardy amerických dolarů. V roce 2008 byl genom Jamese Watsona přečten za několik měsíců. Nové technologie mohou zvládnout jeden lidský genom během dnů a rychlost čtení se bude dále zvyšovat. Tím také skokově klesá cena na 100 000 dolarů; cílem je dosáhnout ceny 1000 dolarů za genom.
3) Genová exprese je děj, při němž se gen nějak projevuje. Zpravidla jde o syntézu molekul RNA, která pak slouží pro překlad do molekul bílkovin s konkrétními funkcemi.
4) V rodinách s častým výskytem studovaného onemocnění sledujeme, zda postižení jedinci sdílejí určité signální znaky. Typicky to jsou opakované sekvence tzv. mikrosatelitů, rozmístěné po celém genomu. Mikrosatelity jsou značně polymorfní – vyskytují se v populaci v mnoha variantách, které se liší délkou, tj. počtem opakování určitého základního motivu. Ve vytipovaných oblastech potom hledáme varianty (alely) genů s možným kauzálním vztahem k onemocnění.
5) Pro pozitivní selekci svědčí převaha mutací DNA, které vedou k záměně určité aminokyseliny v kódovaném proteinu, nad tzv. synonymními mutacemi, při kterých záměna určitého nukleotidu (písmene) v DNA zachovává stejný význam tripletového kódu trojice nukleotidů určujících zařazení jedné aminokyseliny do proteinového řetězce. Stejná aminokyselina je totiž kódována zpravidla několika trojicemi nukleotidů – hovoříme o degeneraci genetického kódu.
6) V nedávné studii byl prokázán dramatický vliv těchto dvou aminokyselinových záměn, které se objevily pravděpodobně v době vzniku jazyka u lidí, na regulaci široké škály genů. Do lidských buněčných neuronálních linií bez endogenního FOXP2 byly vloženy konstrukty DNA s lidskou nebo šimpanzí sekvencí genu FOXP2. Celogenomová analýza exprese genů v takto ovlivněných buňkách ukázala, že u 61 genů došlo k významnému zvýšení a u dalších 55 genů naopak ke snížení exprese při porovnání buněk s lidským a šimpanzím FOXP2. Pod pojmem exprese, vyjádření, zde rozumíme relativní množství mRNA přepisované z příslušných genů, které je detegované pomocí specifické vazby na mikrodestičce s kovalentně navázanou sadou sond (krátkých genových úseků) pokrývajících asi 10 000 genů. Pro detekci navázaného množství je vzorek RNA, resp. komplementární DNA, fluorescenčně značen.
7) Zjednodušeně lze říci, že jde o genetické elementy, které se mohou v genomu samy zmnožovat a transponovat na nová místa. Děje se tak kopírováním do RNA a zpětným přepsáním do DNA, která se může začlenit zpět do genomu.
Ke stažení
 článek ve formátu pdf [277,07 kB]
článek ve formátu pdf [277,07 kB]
O autorovi
Jiří Plachý

Doporučujeme

Lidské ucho v počítači 

Deset let gravitačních vln
