









Co je to gen?
Teprve asi 80 let po Mendelových a Miescherových objevech došlo k zasnoubení genu s DNA. Byly to hlavně výsledky Oswalda Averyho a jeho spolupracovníků (1944), kteří svými experimenty přesvědčivě ukázali, že látka přenesená mezi bakteriálními buňkami při procesu transformace vyvolává dědičnou změnu příjemce a je štěpitelná enzymem, který specificky působí pouze na DNA. Nobelova cena se za války neudělovala, a patrně proto se autoři tohoto epochálního objevu nedočkali ocenění. Teprve Alfred Hershey, který s Marthou Chaseovou jejich objev r. 1952 elegantně potvrdil, dostal Nobelovu cenu (1969) mimo jiné i za zjištění, že bakteriofágy T-řady nasednou na povrch buňky Escherichia coli, ale do jejího nitra vyšlou pouze svou DNA a proteinová obálka viru zůstane venku. DNA stačí na zajištění reprodukce plnohodnotných virových částic uvnitř bakteriální buňky, která pro reprodukci poskytne svoji výbavu zajišťující expresi genů „uložených“ v DNA bakteriofága.
 Dvoušroubovicovou strukturu DNA záhy popsali J. Watson s F. Crickem (1953) a současně navrhli způsob replikace této dvouřetězcové molekuly. Základním poznatkem bylo nalezení kódu (zákona komplementarity), podle kterého se při replikaci dvojřetězcové DNA přiřazují do párů báze adenin, thymin, guanin a cytosin. Možné páry jsou A–T, T–A, G–C a C–G (obr. 1). Tímto objevem se otevřela cesta k pochopení molekulární představy o tocích genetické informace (F. Crick, 1958), kterou zachycuje centrální dogma molekulární genetiky (obr. 2). Zatímco semikonzervativní replikace dvojřetězcové DNA či RNA a transkripce DNA na RNA a reverzní transkripce RNA na DNA probíhají podle variant zákona komplementarity (J. Watson a F. Crick, 1953), syntéza polypeptidického řetězce podle vzoru tzv. mRNA probíhá podle zákona o genetickém kódu (M. Niernberg, H. G. Khorana a další, 1965). V obou případech je tudíž pro pochopení funkce genu důležité znát sekvenci nukleotidů, které gen vytvářejí. Schéma procesu genové exprese (obr. 3) ilustruje typický proces exprese genů prokaryotických buněk, které kódují syntézu proteinů. F. Jacob a J. Monod (1961) popsali základní principy regulace transkripce na příkladu lac-operonu bakterie Escherichia coli a charakterizovali tak podrobněji i výstavbu prokaryotických genů, sestavených často do bloků označovaných jako operony. V rámci operonů mají geny společnou regulaci exprese na transkripční úrovni a vedle několika kódujících oblastí mají tedy navíc oblasti regulační, které rozhodují o tom, zda a kdy se gen projeví (exprimuje).
Dvoušroubovicovou strukturu DNA záhy popsali J. Watson s F. Crickem (1953) a současně navrhli způsob replikace této dvouřetězcové molekuly. Základním poznatkem bylo nalezení kódu (zákona komplementarity), podle kterého se při replikaci dvojřetězcové DNA přiřazují do párů báze adenin, thymin, guanin a cytosin. Možné páry jsou A–T, T–A, G–C a C–G (obr. 1). Tímto objevem se otevřela cesta k pochopení molekulární představy o tocích genetické informace (F. Crick, 1958), kterou zachycuje centrální dogma molekulární genetiky (obr. 2). Zatímco semikonzervativní replikace dvojřetězcové DNA či RNA a transkripce DNA na RNA a reverzní transkripce RNA na DNA probíhají podle variant zákona komplementarity (J. Watson a F. Crick, 1953), syntéza polypeptidického řetězce podle vzoru tzv. mRNA probíhá podle zákona o genetickém kódu (M. Niernberg, H. G. Khorana a další, 1965). V obou případech je tudíž pro pochopení funkce genu důležité znát sekvenci nukleotidů, které gen vytvářejí. Schéma procesu genové exprese (obr. 3) ilustruje typický proces exprese genů prokaryotických buněk, které kódují syntézu proteinů. F. Jacob a J. Monod (1961) popsali základní principy regulace transkripce na příkladu lac-operonu bakterie Escherichia coli a charakterizovali tak podrobněji i výstavbu prokaryotických genů, sestavených často do bloků označovaných jako operony. V rámci operonů mají geny společnou regulaci exprese na transkripční úrovni a vedle několika kódujících oblastí mají tedy navíc oblasti regulační, které rozhodují o tom, zda a kdy se gen projeví (exprimuje).
 Objev P. Sharpa a R. Robertse (1977) týkající se nespojitých genů běžných v eukaryotických organismech vedl k vytvoření dosti odlišného schématu exprese pro buňky eukaryotické (obr. 4). Tyto a další výsledky, které se staly základem nového oboru, molekulární genetiky, respektive molekulární biologie vedly, k sérii podobných definic genů.
Objev P. Sharpa a R. Robertse (1977) týkající se nespojitých genů běžných v eukaryotických organismech vedl k vytvoření dosti odlišného schématu exprese pro buňky eukaryotické (obr. 4). Tyto a další výsledky, které se staly základem nového oboru, molekulární genetiky, respektive molekulární biologie vedly, k sérii podobných definic genů.
Zvláště významnou roli v období formulace centrálního dogmatu, nejen u nás, ale na celém světě, sehrála Watsonova učebnice Molekulární biologie genu, která v českém překladu třetího vydání z roku 1976 vyšla u nás r. 1982. Ve slovníku této publikace se objevuje následující definice: Gen je úsek chromozomu, který kóduje funkční produkt, přičemž funkčním produktem je míněna RNA nebo její translační produkt, polypeptid. Tato definice je z dnešního hlediska, myslím, dosti problematická hlavně proto, že používá termín chromozom, což je nadmolekulární útvar obsahující kromě kódující DNA i další složky, a navíc existují např. plazmidové nebo mitochondriální DNA, které se neoznačují jako chromozomy, a přesto obsahují geny. Kromě toho není brána v potaz RNA, která u některých virů a i v některých buňkách také může nést exprimovatelné geny. O něco lepší je mnohem pozdější definice uvedená např. v novém vydání Biologie pro gymnázia (J. Jelínek, V. Zicháček, Nakladatelství Olomouc, 2007): Gen (vloha) jako jednotka odpovědná za vznik dědičné vlastnosti je z hlediska molekulární biologie úsekem DNA, který svým pořadím nukleotidů určuje pořadí aminokyselin v určité bílkovině nebo pořadí nukleotidů v molekulách RNA. I tato definice však opomíjí RNA-geny, což se dá ovšem snadno napravit: Gen… je úsekem DNA, popřípadě vzácně také RNA…
Společnou slabinou těchto definic je, že i po úpravě zůstává obsah pojmu gen zúžen pouze na kódující sekvenci. Opomíjejí se sekvence regulační a další funkční sekvence provázející sekvenci kódující, v nichž může vzniknout mutace vedoucí ke změně pozorovatelné vlastnosti (fenotypu) podobně jako po mutaci v kódující oblasti. Tento problém částečně řeší definice z pulikace Heleny Pearsonové3) z r. 2006 (s podobnými definicemi se setkáváme i v řadě encyklopedií): Gen je lokalizovatelná oblast genomové sekvence odpovídající jednotce dědičnosti, která sestává z regulačních oblastí, transkribované oblasti a dalších funkčních sekvencí. Oblast kódující protein může být vynechána, protože je vlastně již obsažena v oblasti transkribované. Bohužel, lokalizovatelná oblast genomové sekvence je termín poněkud mlhavý a podobně jako termín úsek DNA se podle výsledků z postgenomové éry ukazuje jako mírně řečeno nepřesný.
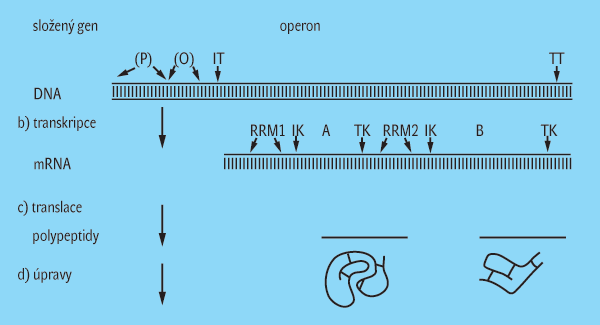 Již ve vysokoškolské učebnici Obecná genetika (J. Nečásek, I. Cetl a spol., 1979), na níž jsem se podílel dodatkem o struktuře a složení nukleových kyselin a proteinů, je upozorněno, že určité části molekul DNA mohou mít i jiné než výše uvedené dědičné funkce spojené s produkcí RNA či proteinů. Definice genů z období dovršení centrálního dogmatu tyto sekvence mezi geny nezahrnují. Tato připomínka je, myslím, platná dodnes. Obecně lze říci, že definice z této etapy oproti původnímu, mendelovskému pojetí genu jako určité jednotky dědičnosti projevující se zjevnou děděnou vlastností (fenotypem) zužuje obsah pojmu gen na sekvence kódující, nebo v lepším případě ke kódujícím sekvencím přidružuje ještě nekódující sekvence, jež kódující úsek DNA či RNA obklopují a mají nějaký funkční vztah, obvykle zajišťují exprese genetické informace v něm zaznamenané. Navíc se ukázalo, že to, co platí o strukturním uspořádání a expresi genů např. v buňkách prokaryotických, neplatí v plné míře v buňkách eukaryotických. Zatímco prokaryotické buňky mají často kódující sekvence sdružované do skupin (operonů), eukaryotické geny se v této etapě jeví spíše jako oddělené (jednotlivé), přičemž čím je organismus komplexnější, tím se častěji setkáváme s geny, které jsou v kódující oblasti nespojité a jejich primárně syntetizovaná RNA musí být sestřihem upravena na funkční formu.
Již ve vysokoškolské učebnici Obecná genetika (J. Nečásek, I. Cetl a spol., 1979), na níž jsem se podílel dodatkem o struktuře a složení nukleových kyselin a proteinů, je upozorněno, že určité části molekul DNA mohou mít i jiné než výše uvedené dědičné funkce spojené s produkcí RNA či proteinů. Definice genů z období dovršení centrálního dogmatu tyto sekvence mezi geny nezahrnují. Tato připomínka je, myslím, platná dodnes. Obecně lze říci, že definice z této etapy oproti původnímu, mendelovskému pojetí genu jako určité jednotky dědičnosti projevující se zjevnou děděnou vlastností (fenotypem) zužuje obsah pojmu gen na sekvence kódující, nebo v lepším případě ke kódujícím sekvencím přidružuje ještě nekódující sekvence, jež kódující úsek DNA či RNA obklopují a mají nějaký funkční vztah, obvykle zajišťují exprese genetické informace v něm zaznamenané. Navíc se ukázalo, že to, co platí o strukturním uspořádání a expresi genů např. v buňkách prokaryotických, neplatí v plné míře v buňkách eukaryotických. Zatímco prokaryotické buňky mají často kódující sekvence sdružované do skupin (operonů), eukaryotické geny se v této etapě jeví spíše jako oddělené (jednotlivé), přičemž čím je organismus komplexnější, tím se častěji setkáváme s geny, které jsou v kódující oblasti nespojité a jejich primárně syntetizovaná RNA musí být sestřihem upravena na funkční formu.
 Genomové projekty otevřely další etapu. V souvislosti s nimi (ale částečně již i dříve) se začaly hromadit údaje o genech, které kódují proteiny nebo RNA, ale jejich výstavba neodpovídá jednoduchým schématům uvedeným na obr. 3 a 4, a tudíž vznikla potřeba stávající definice upravit tak, aby byly s těmito poznatky v souladu. Kromě toho se hromadí také stále nová zjištění o možné účasti i jiných způsobů záznamu děděné informace než prostřednictvím sekvence čtyř nukleotidů v DNA (popřípadě v RNA).
Genomové projekty otevřely další etapu. V souvislosti s nimi (ale částečně již i dříve) se začaly hromadit údaje o genech, které kódují proteiny nebo RNA, ale jejich výstavba neodpovídá jednoduchým schématům uvedeným na obr. 3 a 4, a tudíž vznikla potřeba stávající definice upravit tak, aby byly s těmito poznatky v souladu. Kromě toho se hromadí také stále nová zjištění o možné účasti i jiných způsobů záznamu děděné informace než prostřednictvím sekvence čtyř nukleotidů v DNA (popřípadě v RNA).
Protože hlavní proud v genomové, resp. postgenomové éře se věnuje charakterizaci a porovnávání různých forem genů kódujících proteiny a RNA, pokusím se v dalším pokračování nejprve shrnout nejdůležitější problémy, které nové poznatky o kódujících genech způsobily, a prohloubily tak neuspokojivý stav definice genu. Teprve v dalších pokračováních se budu věnovat dědičnosti nezaložené na sekvencích kódujících proteiny či RNA, které se v této fázi vývoje genu z naší pozornosti vytratily.
Poznámky
1) Proteiny, které usnadňují pasivní průstup malých molekul buněčnou membránou.
2) Cistron je oblast genu, která kóduje jeden polypeptid. U prokaryot se vyskytuje polycistronická mRNA, u eukaryot monocistronická mRNA.
3) Nature 441, 398–401, 2006; doi: 10.1038/441398a
Ke stažení
 článek ve formátu pdf [474,56 kB]
článek ve formátu pdf [474,56 kB]
O autorovi
Vladimír Vondrejs

Doporučujeme

Ideologie v mapách, mapy v rukách ideologů 

Pátranie po pôvode SARS-CoV-2 pokračuje
